|
ТРАНСКРИПЦИЯ (от
лат. transcriptio, букв-переписывание), биосинтез РНК на матрице ДНК; первая
стадия реализации генетич. информации, в ходе к-рой нуклеотидная последовательность
ДНК считывается в виде нуклеотидной последовательности РНК (см. Генетический
код). В основе этого процесса лежит принцип комплементарного спаривания
пуриновых и пиримидиновых оснований (см. Компле-ментарность). Т.
осуществляется с участием фермента РНК-полимеразы, использующей в качестве
субстратов рибонук-леозидтрифосфаты. Кроме того, в транскрипции участвует большое
число вспомогат. белков, регулирующих работу РНК-полимеразы.
Т. происходит на участках
ДНК, наз. единицами Т. или трапскриптонами. В начале и конце транскрилтона расположены
специфич. нуклеотидные последовательности -соотв. промотор и терминатор. Существование
множества транскриптонов обеспечивает возможность независимого считывания разных
генов, их индивидуального включения и выключения. У животных, растений и др.
эукариот в состав транскриптона, как правило, входит один ген. Транс-криптоны
бактерий обычно наз. оперонами; мн. из них содержат по неск. генов, обычно функционально
связанных (напр., кодирующих неск. ферментов, участвующих в синтезе той или
иной аминокислоты).
Процесс синтеза РНК можно
разделить на четыре основные стадии: 1) связывание РНК-полимеразы с промотором,
2) начало синтеза цепи РНК (инициация), 3) рост цепи РНК (элонгация), 4) завершение
синтеза цепи РНК (терминация).
Связывание РНК-полимеразы
с промотором включает по крайней мере два этапа. На первом РНК-полимераза образует
с промотором закрытый комплекс, в к-ром ДНК сохраняет двухспиральную структуру,
а РНК-полимераза еще не способна начать синтез РНК. На втором закрытый комплекс
превращается в открытый, в к-ром РНК-полимераза расплетает примерно один виток
двойной спирали ДНК в районе стартовой точки-нуклеотида, с к-рого начинается
комплементарное копирование матрицы.
При наличии субстратов
РНК-полимераза в открытом комплексе осуществляет инициацию. Первый нуклеотид
(обычно это аденозин- или гуанозинтрифосфат) входит в состав цепи целиком, а
последующие присоединяются к группе 3'-ОН предыдущего нуклеотида с образованием
фос-фодиэфирной связи и освобождением пирофосфата (см. Нуклеиновые кислоты).
На стадии инициации образующаяся РНК связана с матрицей и ферментом непрочно
и может отделиться от комплекса. В этом случае РНК-полимераза, не покидая промотора,
снова инициирует РНК (такой синтез коротких рибонуклеотидов наз. абортивным).
Стадия инициации завершается,
когда цепь РНК достигает критич. длины (от 3 до 9 нуклеотидов на разных промоторах);
при этом от РНК-полимеразы отделяется s-субъединица.
Считают, что в процессе
элонгации примерно 13 нуклеотидов РНК образуют гибридную спираль с матричной
нитью расплетенной ДНК (всего на этой стадии в ДНК расплетено примерно 18 нуклеотидов).
По мере движения РНК-полимеразы по матрице впереди нее происходит расплетание,
а позади восстановление двойной спирали ДНК. Одновременно происходит вытеснение
очередного звена растущей цепи РНК из комплекса с матрицей.
Цепь РНК растет в направлении
5' : 3' по мере продвижения РНК-полимеразы по цепи ДНК в направлении от
3'-конца к 5'-концу. Средняя скорость роста цепи РНК у бактерии Escherichia
coli (E. coli) составляет 40-45 рибонуклеотидов в секунду. В процессе удлинения
цепи РНК фермент движется по ДНК с непостоянной скоростью. В нек-рых участках
матрицы происходят длительные задержки в его продвижении, т. наз. паузы (нек-рые
стадии Т. показаны на рис.).
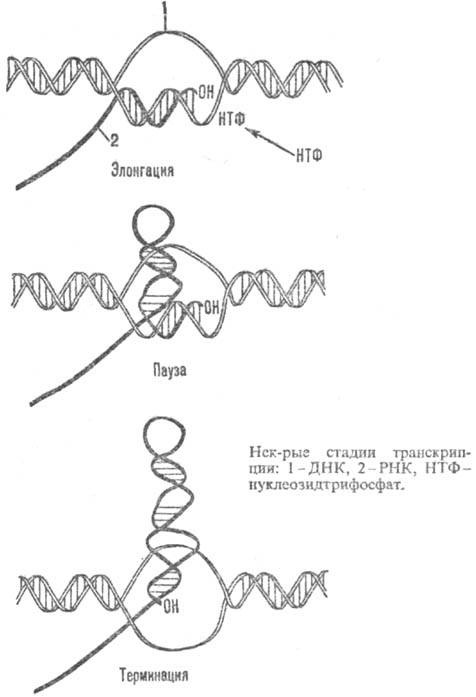
На стадии злонгации в состав
транскрибирующего комплекса входит ряд дополнит. белков, от к-рых зависит протекание
завершающей стадии транскрипции -терминации. Один из таких белков, кодируемых
геном nusA E. coli, занимает в РНК-полимеразе место s-субъединицы.
Др. бактериальный фактор терминации r взаимод. с РНК.
Терминация Т., как правило,
происходит в строго определенных участках матрицы - терминаторах, в к-рых от
матрицы отделяются РНК и РНК-полимераза; последняя, объединившись со свободной
s-субъединицей, может вступить в следующий цикл Т. В терминаторах, для
узнавания к-рых РНК-полимеразе не требуется фактора р, нуклеотидная последовательность
характеризуется двумя особенностями: по ходу Т. перед точкой терминации расположен
участок, богатый парами dG-dC (дезоксигуанозин-дезоксицити-дин), а затем участок,
состоящий из 4-8 расположенных подряд остатков дезоксиадениловой к-ты. Предполагают,
что после прохождения РНК-полимеразой участка, богатого dG-dC, в РНК возникает
шпилька, к-рая препятствует продвижению фермента и разрушает часть спирали РНК-ДНК
транскрибирующего
комплекса. Оставшаяся часть гибридной спирали, включающая концевую полиуридиловую
последовательность РНК, легко плавится (разрушается) ввиду крайней нестабильности
комплементарной пары уридин-дезоксиаденозин, что и приводит к освобождению РНК.
Мн. терминаторы узнаются
РНК-полимеразой только с помощью белковых факторов терминации. Из них наиб.
изучен фактор r E. coli-олигомерный белок с мол. м. 46 тыс. Фактор
r присоединяется к определенным участкам синтезируемой РНК (не имеющим
протяженных двухспи-ральных структур) до того, как РНК-полимераза достигает
терминатора. Предполагается, что фактор r передвигается вдоль РНК вслед
за РНК-полимеразой, используя для этого энергию гидролиза нуклеозидтрифосфатов,
и способствует диссоциации гибрида РНК с матричной нитью ДНК.
Скорость Т. разл. генов
может отличаться в тысячи раз; в столь же больших пределах может изменяться
скорость Т. одного и того же гена в разных тканях многоклеточного организма
или в одной клетке в зависимости от изменяющихся внеш. условий или внутр. программы.
На стадии инициации регуляция Т. осуществляется благодаря наличию особых белков-регуляторов
(см. Регуляторные белки), способных присоединяться к определенным
участкам ДНК и тем самым препятствовать или помогать РНК-полимеразе инициировать
синтез РНК на промоторе.
У прокариот регуляция Т.
часто осуществляется на стадии терминации в особых терминаторах (называемых
аттенюаторами), расположенных в начале или внутри оперонов.
Существует также обратная
Т.-синтез ДНК на матрице РНК. Такой синтез осуществляется у ретровирусов (семейство
РНК-содержащих вирусов) с участием фермента ревер-тазы (обратная транскриптаза).
В ходе обратной Т. образуется вначале гибрид РНК-ДНК, к-рый реплицирует под
действием ДНК-зависимой ДНК-полимеразы (см. Полиде-зоксирибонуклеотид-синтетазы)
с образованием двухцепо-чечной спирали ДНК. Последняя также подвержена репликации
и способна включаться в геном инфицированной клетки и служить там матрицей для
вирусной РНК. Т. обр., поток генетич. информации у ретровирусов направлен от
РНК к ДНК и затем обратно к РНК.
РНК-полимеразу открыли
С. Вейс, Ж. Гурвиц и О. Стивене в 1960; ими же установлено ее значение в синтезе
РНК. Концепцию транскриптона (оперона) сформулировали Ф. Жакоб и Ж. Моно в 1961.
X. Темин и Д. Балтимор в 1970 открыли обратную транскриптазу и механизм синтеза
ДНК на РНК-матрице.
===
Исп. литература для статьи «ТРАНСКРИПЦИЯ»: Пташне М.,
Переключение генов: регуляция генной активности и фаг l, пер. с англ.,
М., 1988; Молекулярная биология. Структура и биосинтез нуклеиновых кислот, М.,
1990. В. Г. Никифоров.
Страница «ТРАНСКРИПЦИЯ» подготовлена по материалам химической энциклопедии.
|



